Иммунологическая память: общая характеристика
Иммунологическая память - это способность иммунной системы отвечать более быстро и эффективно на антиген (патоген), с которым у организма был предварительный контакт.
Такая память обеспечивается предсуществующими антигенспецифическими клонами как В-клеток , так и Т-клеток , которые функционально более активны в результате прошедшей первичной адаптации к определенному антигену.
Пока неясно, устанавливается ли память в результате формирования долгоживущих специализированных клеток памяти или же память отражает собой процесс рестимуляции лимфоцитов постоянно присутствующим антигеном, попавшим в организм при первичной иммунизации.
Клетки иммунологической памяти
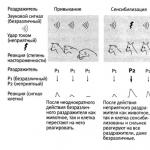
Вторичный иммунный ответ характеризуется более быстрой и эффективной продукцией антител .Интенсивность ответа, осуществляемого популяцией примированных B-лимфоцитов, возрастает, главным образом, за счет увеличения клеток, способных воспринимать антигенный стимул ( рис. 2.13-R ). На рисунке схематически представлено образование эффекторных клеток и клеток памяти после первичного контакта с антигеном. Часть потомков антигенреактивных лимфоцитов после устранения инфекции превращается в неделящиеся клетки памяти, а остальные становятся эффекторными клетками клеточного иммунитета. Клеткам памяти требуется меньше времени для того, чтобы активироваться при повторной встрече с антигеном, что соответственно укорачивает интервал, необходимый для возникновения вторичного ответа.
B-клетки иммунологической памяти качественно отличаются от непремированных B- лимфоцитов не только тем, что начинают продуцировать IgG -антитела раньше, но они обычно обладают и более высокоаффинными антигенными рецепторами благодаря селекции в ходе первичного ответа.
T-клетки памяти вряд ли обладают рецепторами повышенной аффинности по сравнению с непримированными T-клетками. Однако T-клетки иммунологической памяти способны реагировать на более низкие дозы антигена, и это позволяет предполагать, что их рецепторный комплекс в целом (включая молекулы адгезии ) функционирует более эффективно.
Таким образом можно считать установленным, что иммунологическая память определяется не только накоплением популяций одинаковых по свойствам клеток; меняются также свойства индивидуальных клеток, о чем свидетельствуют изменения в экспрессии молекул клеточной поверхности и цитокинов.
B-клеточная иммунологическая память
Общая характеристика В-клеток при вторичном ответе, которая собственно и определяет В-клеточную память , включает следующие показатели.1). На порядок увеличивается количество специфических В- клеток, вступающих во вторичный ответ, в сравнении с количеством этих клеток при первичном ответе. Например, отношение антигенспецифических В-клеток к общему содержанию В-клеток в селезенке при первичном иммунном ответе к патогенам составляет приблизительно 1:10000; в то же время при вторичном ответе это отношение равно 1:1000.
2). Сокращается латентный период и раньше достигается максимум продукции антител . Для разных антигенов эти показатели варьируют, однако, в среднем, время латентного периода и достижения пика антител при вторичном ответе уменьшается на 2-4 дня.
3). При первичном ответе доминирует продукция IgM . Вторичный ответ характеризуется преимущественной продукцией IgG .
4). Повышается аффинность антител.
Все эти характерологические признаки В-клеточной памяти закладываются при развитии первичного иммунного ответа. В это время происходит накопление антигенспецифического клона В-клеток, идет процесс его дифференцировки, осуществляется отбор клонов на наибольшую аффинность с помощью .
При вторичном ответе принципиальные события очевидно те же, что и при первичном ответе. Однако в реакцию на антиген вступают уже подготовленные клетки с высокоаффинными антигенраспознающими рецепторами. Возможно, при вторичном ответе идет дополнительное повышение аффинности рецепторов, что определяет еще большее сродство антител к антигену. Это предположение строится на экспериментальных данных по последовательному повышению аффинности антител после первичной, вторичной и третичной иммунизации. Зародышевый центр B-лимфоциты: экспрессия CD и этапы гемопоэза
B-лимфоциты: B-клеточные области
Мозг костный
B-лимфоциты: пролиферация тимус-зависимого клона
Антигены: пути распространения
Плазмацитома мышей Фолликулярная B-клеточная лимфома: ген BCL-2 и дифференцировка
T-клеточная иммунологическая память
Быстрота и напряженность вторичного ответа связаны не только с активностью В-клеток памяти , но и с функциональной подготовленностью Т-клеток - наличием Т-клеток памяти .Т-клетки памяти отличаются от наивных Т-клеток изменением экспрессии функционально значимых рецепторов клеточной поверхности ( табл. 13.7 ).
Особое значение имеют различия по L-селектину , CD44 и CD45RO . Первые два белка участвуют в хоминге Т-клеток в лимфоидные органы и очаги проникновения патогена. CD45RO выступает в качестве передатчика сигнала внутрь клетки при формировании антигенраспознающего комплекса.
Изменение экспрессии рецепторов у Т-клеток памяти существенно отличает их от наивных Т-клеток. При этом следует помнить, что констатация подобных изменений не отвечает на вопрос: образуются ли Т-клетки памяти в результате дивергенции наивных Т-клеток в процессе дифференцировки на армированные эффекторные Т-клетки и Т-клетки памяти или же Т-клетки памяти - долгоживущая субпопуляция армированных Т-клеток.
Иначе, являются ли Т-клетки памяти результатом дивергентного или монофилетического развития? Гиперчувствительность типа IV
CD58
Антигены: роль в поддержании иммунологической памяти
Успешно развившийся специфический иммунитет как заключительный этап антиинфекционной защиты разрешает в итоге конфликт между патогеном и организмом в пользу последнего. Выздоровевший организм характеризуется отсутствием легко выявляемых эффекторных антигенспецифических клеток и антител и наличием клеток памяти .Однако все эти факты еще не говорят о полном освобожденнии от антигенов, которыми обладал возбудитель. При работе с мечеными высокомолекулярными антигенами метка была обнаружена на поверхности фолликулярных дендритных клеток через несколько месяцев после иммунизации. Возможно, некоторые антигены того или иного возбудителя могут сохраняться в виде иммунных комплексов на дендритных клетках. Не исключена вероятность длительной персистенции незначительных количеств вирусов или бактериальных клеток, которым удалось "скрыться" от иммунной элиминации. Примером может служить вирус простого герпеса , длительно пребывающий в нервной ткани. Если возбудители действительно ведут себя именно так, то клонам наивных Т-клеток , покидающих тимус , постоянно предоставляется материал для распознавания и дифференцировки в армированные клоноспецифические Т-клетки, что и создает пул постоянно присутствующих подготовленных эффекторов для ответной реакции на повторное проникновение патогена.
Сфинголипиды: влияние на образование клеток памяти
При специфическом распознавании антигена молекула CD4 повышает авидность комплекса TCR/Ag/МНС II класса, а ко-стимуляция CD4 приводит к развитию синергичного пролиферативного ответа. Дифференцировка CD4 + -клеток в Th1 или Th2 происходит при генетически рестриктированном взаимодействии лимфоцита с антиген-презентирующей клеткой , а также определяется плотностью экспрессии рецепторов CD4, CD28 , MEL-14 и др. на лимфоцитах [ Noel, ea 1996 , Deeths, ea 1997 ]. Минорная субпопуляция CD4 + -клеток при этом экспрессирует фенотип активационно-индуцированных клеток памяти ( CD69 high , CD45RB low , CD44 high , L-селектин и т. д.) [ Muralidhar, ea 1996 ]. Образование клеток памяти на Т-зависимые антигены регулируется фумонизином В1 [ Martinova, ea 1995 ].CD4 (T4, gp59)
CD4 (T4 , gp59 , у мышей L3T4 , рецептор ВИЧ ) - это гликопротеин, молекулярная масса которого равна 55 кДа. Полипептидная цепь состоит из 433 аминокислот. CD4 представляет собой одноцепочечную молекулу, состоящую из четырех иммуноглобулинподобных доменов ( рис. 3.17 ). Домены D1 и D2, а также D3 и D4 образуют между собой парные, плотноупакованные, жесткие структуры. Эти пары соединены гибким шарнирным участком. Хвостовая часть молекулы CD4 имеет достаточную длину для взаимодействия с цитоплазматическими белками-трансдукторами. На клеточной поверхности ТКР и CD4 представлены независимо друг от друга. Их встреча происходит в процессе формирования ответа на антиген. После распознавания ТКР антигенного комплекса происходит взаимодействие CD4 с молекулой II класса МНС . Реакция взаимодействия осуществляется между бета2-доменом молекулы МНС и первым доменом CD4. Предполагается также слабое включение во взаимодействие и второго D2-домена.CD4 - представитель суперсемейства Ig , содержащий во внеклеточной части 4 домена. Ig-подобный характер первых двух с N-конца доменов подтвержден с помощью рентгеноструктурного анализа. Домены 3 и 4 гомологичны доменам 1 и 2 CD2 . 6 остатков Cys молекулы формируют три дисульфидные связи. Трансмембранный участок CD4 гомологичен (48%) трансмембранному домену продуктов MHC класса II . Цитоплазматический домен CD4 включает 40 аминокислотных остатка и содержит четыре сайта фосфорилирования. CD4 мышей, крыс, кроликов имеют аналогичное строение и высокую гомологию с CD4 человека (более 50%), особенно в цитоплазматическом участке. В N-концевой части молекулы содержится участок, обладающий сродством к молекуле gp120 ВИЧ .
ФУНКЦИИ. CD4 идентифицирован на поверхности Т-лимфоцитов с помощью моноклональных антител (ОКТ4) в 1979 г. как маркер Т-хелперов . CD4 содержится на поверхности кортикальных тимоцитов , части зрелых периферических Т-лимфоцитов (40-50% - почти исключительно T-хелперов), он обнаруживается также на моноцитах , некоторых клетках головного мозга . На мембране кортикальных тимоцитов CD4 сосуществует с CD8 , тогда как на зрелых Т-клетках экспрессируется CD4 или CD8.
Функция CD4 обусловлена в первую очередь его способностью связываться с молекулами MHC класса II . В связывании антигенов MHC класса II принимают участие два наружных домена CD4 и неполиморфная часть молекулы MHC. Связывание CD4 с антигенами MHC класса II не только обуславливает адгезию CD4плюс Т-хелперов к MHC-IIплюс макрофагам , но и значительно (100-кратно) повышает сродство Т-клеточного рецептора TcR (с которым CD4 необратимо связывается) к комплексу антигена с продуктами MHC класса II. В свою очередь, при связывании TcR-CD3 с антигенным пептидом между CD4 и рецептором формируется (при участии дельта-цепи CD3 ) физический контакт, облегчающий распознавание комплекса антиген- продукт MHC.
и т.д.................
Под иммунной памятью понимают способность организма давать ускоренную иммунологическую реакцию на повторное введение антигена. После первичного ответа на антиген в организме образуется определенное количество долгоживущих клеток памяти, сохраняющих информацию об антигене. При повторном введении антигена в организм клетки памяти обусловливают вторичный иммунный ответ. Основа вторичного ответа та же, что и первичного, однако антителообразование при нем происходит быстрее и более интенсивно, синтезируется преимущественно IgG, аффинитет антител выше, чем при первичном.
Иммунологическая память свойственна Т- и В-лимфоцитам. Так как память к различным антигенам хранят различные клоны лимфоидных клеток, это позволяет лимфоидной системе приобретать новую информацию, не утрачивая предшествующей.
В некоторых случаях возможна ситуация, когда макроорганизм не способен по той или иной причине отвечать на определенные АГ. Такая неотвечаемость получила название имунологической толерантности (толерантность – терпимость, неотвечаемость). Это явление было открыто П.Медаваром на мышах. Оказалось, что если эмбрионам белых мышей ввести клетки селезенки других линий мышей (черных), то взрослые особи, выросшие на этих эмбрионах, не отторгали трансплантанты кожи черных мышей, т.е. становились к ним толерантными. Обычные мыши отторгали такие аллогенные трансплантанты. Аналогичные опыты провел М.Гашек на разных породах кур. В результате экспериментов оказалось, что врожденная толерантность к антигену (толерогену) возникает, когда происходит внутриутробный контакт организма с этим антигеном. В этом случае организм после рождения будет воспринимать данный АГ как “свое”. В настоящее время такая толерантность объясняется тем, что в эмбриогенезе происходит гибель клонов-предшественников Т-лимфоцитов, способных взаимодействовать с толерогеном.
Кроме врожденной, существует также и приобретенная толерантность. Чаще всего это обратимый процесс. Приобретенная толерантность бывает 2-х видов: высокодозовая и низкодозовая. Высокодозовая толерантность возникает при попадании в организм большиз доз толерогена, особенно введенного на фоне подавления иммунитета (облучение, применение иммунодепрессантов). Такое большое количество АГ вызывает гибель реактивных к нему лимфоцитов. Низкодозовая толерантность возникает при введении малых доз определенных АГ. Считается, что в данном случае она опосредована активацией клеток-супрессоров, подавляющих иммунную реакцию. В целом же в настоящее время оба механизма поддержания толерантности (деления клонов и их супрессия) рассматриваются как взаимодополняющие.
Идиотип-антиидиотипическое взаимодействие лежит в основе теории иммунной сети, предложенной Н.К.Ерне (1974 г.) как механизма регуляции функционирования иммунной системы. Сущность его заключается в следующем. К одному и тому же АГ антитела синтезируются различными клонами лимфоцитов. Такие АТ (или, что равнозначно Т-клеточные рецепторы) будут несколько отличаться по строению друг от друга. В активном центре таких АТ или рецепторов находятся уникальные антигенные детерминанты, присущие только данному клону лимфоцитов и отличающие его от любых других. Они получили название идиотипов. Сам АГ-связывающий участок АТ был назван паратоном. Совокупность всех идиотипов данного АТ наз. идиотипом. При развертывании иммунного ответа первоначально синтезируется АТ первого поколения, направленные к данному АГ. Они получили название идиотипических антител (несущих идиотип). К их активным центрам, в свою очередь, впоследствии вырабатываются АТ второго поколения – антиидиотипические. Они блокируют синтез идиотипических АТ. Так осуществляется естественное затухание иммунного ответа, снижающее вероятность развития аутоиммунных процессов.
Иммунологическая память - это способность иммунной системы отвечать более быстро и эффективно на антиген (патоген), с которым у организма был предварительный контакт.
Такая память обеспечивается предсуществующими антигенспецифическими клонами как В-клеток, так и Т-клеток, которые функционально более активны в результате прошедшей первичной адаптации к определённому антигену.
Такая память обеспечивается предсуществующими антигенспецифическими клонами как В-клеток, так и Т-клеток, которые функционально более активны в результате прошедшей первичной адаптации к определенному антигену.
В результате первой встречи запрограммированного лимфоцита с определенным антигеном образуются две категории клеток: эффекторные, которые немедленно выполняют специфическую функцию - секретируют антитела или реализуют клеточные имунные реакции, и клетки памяти, которые циркулируют длительное время. При повторном поступлении данного антигена они быстро превращаются в лимфоциты-эффекторы, которые вступают в реакцию с антигеном. При каждом делении запрограммированного лимфоцита после его встречи с антигеном количество клеток памяти увеличивается.
Клеткам памяти требуется меньше времени для того, чтобы активироваться при повторной встрече с антигеном, что соответственно укорачивает интервал, необходимый для возникновения вторичного ответа.
B-клетки иммунологической памяти качественно отличаются от непремированных B-лимфоцитов не только тем, что начинают продуцировать IgG -антитела раньше, но они обычно обладают и более высокоаффинными антигенными рецепторами благодаря селекции в ходе первичного ответа.
T-клетки памяти вряд ли обладают рецепторами повышенной аффинности по сравнению с непримированными T-клетками. Однако T-клетки иммунологической памяти способны реагировать на более низкие дозы антигена, и это позволяет предполагать, что их рецепторный комплекс в целом (включая молекулы адгезии) функционирует более эффективно.
Вакцины живые, убитые, химические, анатоксины, синтетические вакцины. Современные рекомбинантные вакцины. Принципы поучения каждого вида вакцин, механизмы создаваемого иммунитета. Адъюванты в вакцинах.
Живые вакцины содержат жизнеспособные штаммы патогенных микробов, ослабленные до степени, исключающей возникновение заболевания, но полностью сохранившие антигенные и иммуногенные свойства. Это аттенуированные в естественных или искусственных условиях штаммы микроорганизмов. Аттенуированные штаммы вирусов и бактерий получают путем инактивации генов, ответственных за образование факторов вирулентности, или за счет мутаций в генах, неспецифически снижающих эту вирулентность. Вакцинные штаммы микроорганизмов, сохраняя способность размножаться, вызывают развитие бессимптомной вакцинальной инфекции. Реакцию организма на введение живой вакцины расценивают не как болезнь, а как вакцинальный процесс. Вакцинальный процесс продолжается несколько недель и приводит к формированию иммунитета к патогенным штаммам микроорганизмов.
Живые вакцины имеют ряд преимуществ перед убитыми и химическими вакцинами. Живые вакцины создают прочный и длительный иммунитет, по напряженности приближающийся к постинфекционному. Для создания прочного иммунитета во многих случаях достаточно одного введения вакцины, причем, такие вакцины могут вводиться в организм достаточно простым методом – например, скарификационным или пероральным. Живые вакцины используют для профилактики таких заболеваний, как полиомиелит, корь, паротит, грипп, чума, туберкулез, бруцеллез, сибирская язва.
Для получения аттенуированных штаммов микроорганизмов используют следующие методы.
1. Культивирование высокопатогенных для человека штаммов путем последовательных пассажей через культуры клеток или организм животных, либо путем воздействия во время роста и размножения микробов физическими и химическими факторами. В качестве таких факторов могут быть использованы необычная температура, неблагоприятные для роста питательные среды, ультрафиолетовое облучение, формалин и др. факторы. Подобным образом были получены вакцинные штаммы возбудителя сибирской язвы, туберкулеза.
2). Адаптация к новому хозяину – пассирование возбудителя на невосприимчивых животных. Путем длительного пассирования через мозг кролика вируса уличного бешенства Пастер получил фиксированный вирус бешенства, который был максимально вирулентен для кролика и минимально вирулентен для человека, собак, сельскохозяйственных животных.
2) Выявление и селекция штаммов микроорганизмов, утративших в естественных условиях вирулентность для человека (вирус осповакцины).
3) Создание вакцинных штаммов микроорганизмов с помощью методов генной инженерии путем рекомбинации геномов вирулентного и невирулентного штаммов.
Недостатки живых вакцин:
Остаточная вирулентность
Высокая реактогенность
Генетическая нестабильность – ревертирование к дикому типу, т.е. восстановление вирулентных свойств
Способность вызывать тяжелые осложнения, в том числе эецефалиты и генерализацию вакцинного процесса.
Убитые вакцины, способы получения, использование для профилактики и терапии инфекционных заболеваний, создаваемый иммунитет, примеры;
Убитые (корпускулярные) вакцины содержат взвесь цельных микробных клеток, инактивированных физическими и химическими методами. Микробная клетка сохраняет антигенные свойства, но лишаются жизнеспособности. Для инактивации используют нагревание, ультрафиолетовое облучение, формалин, фенол, спирт, ацетон, мертиолят и др. Убитые вакцины обладают более низкой эффективностью по сравнению с живыми вакцинами, но при повторном введении создают достаточно стойкий иммунитет. Вводятся парентерально. Корпускулярные вакцины применяют для профилактики таких заболеваний, как брюшной тиф, холера, коклюш и др.
- химические (субъединичные) вакцины, способы получения, использование, создаваемый иммунитет, примеры;
Химические (субъединичные) вакцины содержат специфические антигены, извлеченные из микробной клетки с помощью химических веществ. Из микробных клеток извлекают протективные антигены, представляющие собой иммунологически активные вещества, способные при введении в организм обеспечивать формирование специфического иммунитета. Протективные антигены находятся либо на поверхности микробных клеток, либо в клеточной стенке, либо на клеточной мембране. По химической структуре они представляют собой либо гликопротеиды, либо белково- полисахаридно-липидные комплексы. Извлечение антигенов из микробных клеток осуществляется различными способами: экстрагированием кислотой , гидроксиламином, осаждением антигенов спиртом, сернокислым аммонием, фракционированием. Полученная таким путем вакцина содержит специфические антигены в высокой концентрации и не содержит балластных и токсических субстанций. Химические вакцины обладают низкой иммуногенностью, поэтому вводятся с адъювантами. Адъюванты - это вещества, которые сами по себе не обладают антигенными свойствами, но при введениии с каким-либо антигеном усиливают иммунный ответ на данный антиген. Такие вакцины используются для профилактики менингококковой инфекции, холеры и др.
Расщепленные (сплит) вакцины, их характеристика, применений для профилактики инфекционных заболеваний, примеры;
Расщепленные вакцины готовятся обычно из вирусов и содержат отдельные антигены вирусной
частицы. Они, также, как и химические, обладают низкой иммуногенностью, поэтому вводятся с
адьювантом. Примером подобной вакцины является вакцина против гриппа.
- искусственные вакцины, их разновидности, характеристика, применение, примеры;
- рекомбинантные вакцины, получение, применение, примеры.
Рекомбинантные вакцины - это вакцины, разработанные на основе генно-инженерных методов. Принцип создания генно-инженерных вакцин включает выделение природных генов антигенов или их активных фрагментов, встройку этих генов в геном простых биологических объектов (бактерии, например, кишечная палочка, дрожжи, крупные вирусы). Необходимые для приготовления вакцины антигены получают при культивировании биологического объекта, который является продуцентом антигена. Подобная вакцина используется для профилактики гепатита В.
Препараты, содержащие антитела (гипериммунная плазма, антитоксические, антимикробные сыворотки, гамма-глобулины и иммуноглобулины), их характеристика, получение, титрование. Серотерапия и серопрофилактика.
Б) препараты, содержащие антитела:
Классификация препаратов, содержащих антитела
· Лечебные сыворотки.
· Иммуноглобулины.
· Гамма-глобулины.
· Препараты плазмы.
Различают два источника получения специфических сывороточных препаратов:
1) гипериммунизация животных (гетерологичные сывороточные препараты);
2) вакцинация доноров (гомологичные препараты).
Сыворотки антимикробные и антитоксические, гомологичные и гетерологичные, получение, титрование, очистка от балластных белков, применение, создаваемый иммунитет, примеры;
Антимикробные сыворотки содержат антитела против клеточных антигенов возбудителя. Их получают иммунизацией животных клетками соответствующих возбудителей и дозируют в миллилитрах. Антимикробные сыворотки могут применяться при лечении:
Сибирской язвы;
Стрептококковых инфекций;
Стафилококковой инфекции;
Синегнойной инфекции.
Их назначение определяется тяжестью течения заболевания и, в отличие от антитоксических, не является обязательным. При лечении больных с хроническими, длительно, вяло текущими формами инфекционных заболеваний возникает необходимость стимулировать собственные механизмы специфической зашиты путем введения различных антигенных препаратов и создания активного приобретенного искусственного иммунитета {иммунотерапия антигенными препаратами). Для этих целей используются в основном лечебные вакцины и значительно реже - аутовакцины или стафилококковый анатоксин.
Антитоксические сыворотки содержат антитела против экзотоксинов. Их получают путем гипериммунизации животных (лошадей) анатоксином.
Активность таких сывороток измеряется в АЕ (антитоксических единицах) или ME (международных единицах) - это минимальное количество сыворотки, способное нейтрализовать определенное количество (обычно 100 DLM) токсина для животных определенного вида и определенной массы. В настоящее время в России
антитоксические сыворотки:
Противодифтерийная;
Противостолбнячная;
широко используются следующие
Противогангренозная;
Противоботулиническая.
Применение антитоксических сывороток при лечении соответствующих инфекций обязательно.
Гомологичные сывороточные препараты получают из крови доноров, специально иммунизированных против определенного возбудителя или его токсинов. При введении таких препаратов в организм человека антитела циркулируют в организме несколько дольше, обеспечивая пассивный иммунитет или лечебный эффект в течение 4-5 недель. В настоящее время применяют донорские иммуноглобулины нормальные и специфические и донорскую плазму. Выделение иммунологически активных фракций из донорских сывороток производят с использованием спиртового метода осаждения. Гомологичные иммуноглобулины практически ареактогенны, поэтому реакции анафилактического типа при повторных введениях гомологичных сывороточных препаратов возникают редко.
Для изготовления гетерологичных сывороточных препаратов используют в основном крупных животных лошадей. Лошади обладают высокой иммунологической реактивностью, от них в сравнительно короткий срок можно получить сыворотку, содержащую антитела в высоком титре. Кроме этого, введение лошадиного белка человеку дает наименьшее количество побочных реакций. Животные других видов используются редко. Годные к эксплуатации в возрасте от 3 лет и выше животные подвергаются гипериммунизации, т.е. процессу многократного введения возрастающих доз антигена с целью накопления в крови животных максимального количества антител и поддержания его на достаточном уровне в течение возможно более длительного времени. В период максимального нарастания титра специфических антител в крови животных осуществляют 2-3 кровопускания с интервалом в 2дня. Кровь берут из расчета 1 литр на 50 кг веса лошади из яремной вены в стерильную бутыль, содержащую антикоагулянт. Полученная от лошадей-продуцентов кровь передается в лабораторию для дальнейшей обработки. Плазма отделяется на сепараторах от форменных элементов и дефибринируется раствором хлористого кальция. Использование цельной гетерологичной сыворотки сопровождается аллергическими реакциями в форме сывороточной болезни и анафилаксии. Одним из путей уменьшения побочных реакций сывороточных препаратов, а также повышения их эффективности является их очистка и концентрация. Сыворотку очищают от альбуминов и некоторых глобулинов, которые не относятся к иммунологически активным фракциям сывороточных белков. Иммунологически активными являются псевдоглобулины с электрофоретической подвижностью между гамма- и бета-глобулинами, к этой фракции относятся антитоксические антитела. Также к иммунологически активным фракциям относятся гамма-
глобулины, в эту фракцию входят антибактериальные и антивирусные антитела. Очистка сывороток от балластных белков проводится по методу «Диаферм-3». При использовании этого метода сыворотка очищается путем осаждения под влиянием сернокислого аммония и путем пептического переваривания. Помимо метода «Диаферм 3»,разработаны и другие (Ультраферм, Спиртоферм, иммуносорбцииидр.), имеющие ограниченное применение
Содержание антитоксина в антитоксических сыворотках выражается в международных единицах (ME), принятых ВОЗ. Например, 1 ME противостолбнячной сыворотки соответствует ее минимальному количеству, нейтрализующему 1000 минимальных смертельных доз (DLm) столбнячного токсина для морской свинки массой 350 г. 1 ME противоботулинического антитоксина - наименьшее количество сыворотки, нейтрализующее 10000 DLm ботулинического токсина для мышей массой 20 г. 1 ME противодифтерийной сыворотки соответствует ее минимальному количеству, нейтрализующему 100 DLm дифтерийного токсина для морской свинки массой 250 г.
В препаратах иммуноглобулинов IgG является основным компонентом (до 97%). lgA, IgM, IgD входят в препарат в очень малых количествах. Выпускаются также препараты иммуноглобулинов (IgG), обогащенные IgM и IgA. Активность препарата иммуноглобулина выражается в титре специфических антител, определяемых одной из серологических реакций и указывается в наставлении по применению препарата.
Гетерологичные сывороточные препараты применяют для лечения и профилактики инфекционных заболеваний, вызываемых бактериями, их токсинами, вирусами. Своевременное раннее применение сыворотки может не дать развиться болезни, удлиняется срок инкубации, появившееся заболевание имеет более мягкое течение, снижается смертность.
Существенным недостатком использования гетерологичных сывороточных препаратов является возникновение сенсибилизации организма к чужеродному белку. Как указывают исследователи, к глобулинам сыворотки лошади в России сенсибилизировано более 10% населения. В связи с этим повторное введение гетерологичных сывороточных препаратов может сопровождаться осложнениями в виде различных аллергических реакций, самой грозной из которых является анафилактический шок.
Для выявления чувствительности пациента к лошадиному белку ставят внутрикожную пробу с разведенной 1:100 лошадиной сывороткой, которую специально изготавливают для этой цели. Перед введением лечебной сыворотки пациенту внутрикожно на сгибательную поверхность предплечья вводят 0,1 мл разведенной лошадиной сыворотки и наблюдают за реакцией в течение 20 минут.
Гамма-глобулины и иммуноглобулины, их характеристика, получение, применение для профилактики и терапии инфекционных заболеваний, примеры;
Иммуноглобулинами (гамма-глобулинами) называют очищенные и концентрированные препараты гамма-глобулиновой фракции сывороточных белков, содержащие высокие титры антител. Освобождение от балластных сывороточных белков способствует снижению токсичности и обеспечивает быстрое реагирование и прочное связывание с антигенами. Применение гамма-глобулинов снижает количество аллергических реакций и осложнений, возникающих при введении гетерологичных сывороток. Современная технология получения человеческого иммуноглобулина гарантирует гибель вируса инфекционного гепатита. Основным иммуноглобулином в препаратах гамма-глобулина является IgG. Сыворотки и гамма-глобулины вводят в организм различными путями: подкожно, внутримышечно, внутривенно. Возможно также введение в спинномозговой канал. Пассивный иммунитет возникает через несколько часов и длится до двух недель.
Иммуноглобулин антистафилококковый человеческий. Препарат содержит иммунологически активную белковую фракцию, выделенную из плазмы крови доноров, иммунизированных стафилококковым анатоксином. Активным началом являются антитела к стафилококковому токсину. Создает пассивный антистафилококковый антитоксический иммунитет. Используется для иммунотерапии стафилококковой инфекции.
- препараты плазмы, получение, использование для терапии инфекционных заболеваний, примеры; Антибактериальная плазма.
1). Антипротейная плазма. Препарат содержит антипротейные антитела и получается от доноров,
иммунизированных протейной вакциной. При введении препарата создается пассивный
антибактериальный иммунитет. Используется для иммунотерапии ГВЗ протейной этиологии.
2). Антисинегнойная плазма. Препарат содержит антитела к синегнойной палочке. Получается от
доноров, иммунизированных синегнойной корпускулярной вакциной. При введении препарата
создается пассивный специфический антибактериальный иммунитет. Используется для
иммунотерапии синегнойной инфекции.
Антитоксическая плазма.
1) Плазма антитоксическая антисинегнойная. Препарат содержит антитела к экзотоксину А
синегнойной палочки. Получают от доноров, иммунизированных синегнойным анатоксином. При
введении препарата создается пассивный антитоксический антисинегнойный иммунитет.
Используется для иммунотерапии синегнойной инфекции.
2) Плазма антистафилококковая гипериммунная. Препарат содержит антитела к токсину
стафилококка. Получают от доноров, иммунизированных стафилококковым анатоксином. При
введениии создает пассивный антистафилококковый антитоксический иммунитет. Используется для
иммунотерапии стафилококковой инфекции.
Серотерапия (от лат. serum -- сыворотка и терапия), метод лечения заболеваний человека и животных (преимущественно инфекционных) при помощи иммунных сывороток. Лечебный эффект основан на явлении пассивного иммунитета -- обезвреживании микробов (токсинов) антителами (антитоксинами), содержащимися в сыворотках, которые получают путём гипериммунизации животных (главным образом лошадей). Для серотерапии применяют также очищенные и концентрированные сыворотки -- гамма-глобулины; гетерогенные (полученные из сывороток иммунизированных животных) и гомологичные (полученные из сывороток иммунизированных или переболевших людей).
Серопрофилактика (лат. serum сыворотка + профилактика; син.: сывороточная профилактика,) - метод предупреждения инфекционных болезней путем введения в организм иммунных сывороток или иммуноглобулинов. Используется при заведомом или предполагаемом заражении человека. Наилучший эффект достигается при максимально раннем использовании гамма-глобулина или сыворотки.
В отличие от вакцинации при серопрофилактике в организм вводятся специфические антитела, и следовательно, организм практически немедленно становится в той или иной степени резистентным к определенной инфекции. В отдельных случаях серопрофилактика не предупреждая заболевания, приводит к снижению его тяжести, частоты осложнений и летальности. Вместе с тем серопрофилактика обеспечивает пассивный иммунитет лишь в пределах 2-3 нед. Введение сыворотки, полученной из крови животных, в отдельных случаях может вызвать сывороточную болезнь и такое грозное осложнение, как анафилактический шок.
Для предупреждения сывороточной болезни во всех случаях сыворотку вводят по методу Безредки поэтапно: в первый раз - 0,1 мл, через 30 мин - 0,2 мл и через 1 ч всю дозу.
Серопрофилактику проводят против столбняка, анаэробных инфекций, дифтерии, кори, бешенства, сибирской язвы, ботулизма, клещевого энцефалита и др. При ряде инфекционных болезней с целью серопрофилактики одновременно с сывороточными препаратами используют и другие средства: антибиотики при чуме, анатоксин при столбняке и др.
Сыворотки иммунные применяют при лечении дифтерии (преимущественно в начальной стадии болезни), ботулизма, при укусах ядовитых змей; гамма-глобулины -- при лечении гриппа, сибирской язвы, столбняка, оспы, клещевого энцефалита, лептоспироза, стафилококковых инфекций (особенно вызванных антибиотикоустойчивыми формами микробов) и других заболеваний.
Для предупреждения осложнений серотерапии (анафилактический шок, сывороточная болезнь) сыворотки и гетерогенные гамма-глобулины вводят по специальной методике с предварительной кожной пробой.
Иммунологическая память. При повторной встрече с антигеном организм формирует более активную и быструю иммуннуюреакцию - вторичный иммунный ответ. Этот феномен получил название иммунологической памяти.
Иммунологическая память имеет высокую специфичность к конкретному антигену, распространяется как на гуморальное, так и клеточное звено иммунитета и обусловлена В- и Т-лимфоцитами. Она образуется практически всегда и сохраняется годами и даже десятилетиями. Благодаря ней наш организм надежно защищен от повторных антигенных интервенций.
Существует также ограничение ответов генетически отличных людей, что не дает решения. Низкая иммуногенность, вызванная быстрой деградацией пептидов с помощью пептидаз в сыворотке, может быть скорректирована с помощью модификации пептидов или путем их включения в композицию с контролируемым высвобождением.
Могут ли пептидные вакцины использоваться в терапии рака?
Некоторые мутации могут приводить к последовательности, которая распознается Т-лимфоцитами. Другие, такие как мутации р53, вызывают значительно повышенную экспрессию белка из-за структурных изменений , которые препятствуют его деградации. Супер-экспрессия вызывает появление обычно тихих эпитопов. Это способствует знаниям, необходимым для производства конкретных вакцин против мутированных или сверхэкспрессированных последовательностей онкобелков.
На сегодняшний день рассматривают два наиболее вероятных механизма формирования иммунологической памяти. Один из них предполагает длительное сохранение антигена в организме. Этому имеется множество примеров: инкапсулированный возбудитель туберкулеза, персистирующие вирусы кори, полиомиелита, ветряной оспы и некоторые другие патогены длительное время, иногда всю жизнь, сохраняются в организме, поддерживая в напряжении иммунную систему. Вероятно также наличие долгоживущих дендритных АПК, способных длительно сохранять и презентировать антиген.
Такая терапия не используется у людей, но эксперименты с крысами пришли к выводу, что пептидная вакцина, вводимая адъювантом, может вызывать защитный иммунный ответ против опухолевых клеток , имеющих гомологичную мутацию последовательности, используемой для производства вакцины. Рекомбинантная векторная вакцина.
Для создания рекомбинантных вакцин используются несколько различных организмов , таких как бактерии сальмонеллы и вирусы, такие как вакциния и аденовирус. Акцент будет сделан здесь на вакцину и технологию вакцинации на основе аденовирусов. Это выгодно тем, что они очень эффективны при активации гуморального и клеточного иммунного ответа, часто требуя только одного применения. С другой стороны, существуют такие риски, как конверсия вставленных вирусных генов в вирулентность или рекомбинацию с вирусами дикого типа и возможное вмешательство в ранее существовавший иммунитет к вакцинному вектору.
Другой механизм предусматривает, что в процессе развития в организме продуктивного иммунного ответа часть антигенореактивных Т- или В-лимфоцитов дифференцируется в малые покоящиеся клетки, или клетки иммунологической памяти. Эти клетки отличаются высокой специфичностью к конкретной антигенной детерминанте и большой продолжительностью жизни (до 10 лет и более). Они активно рециркулируют в организме, распределяясь в тканях и органах, но постоянно возвращаются в места своего происхождения за счет хоминговых рецепторов. Это обеспечивает постоянную готовность иммунной системы реагировать на повторный контакт с антигеном по вторичному типу.
Эффективность вакцины от вакцинии доказана с помощью экспериментов с вирусом бешенства. Животные, иммунизированные этой вакциной, защищены от летальных доз вируса бешенства. Иммунитет был получен либо с системной, либо с оральной инокуляцией. Его нельзя использовать ни у людей, ни у животных, которые вступают в контакт с ними, потому что у них есть небольшая вероятность обращения к вирулентности.
Он имеет как преимущества, так и высокую эффективность, длительный период воздействия антигена и очень некомпетентность репликации, которая предотвращает нежелательную пролиферацию вирусного вектора. В основном из-за аспекта репликации-некомпетентности эта вакцина была предметом изучения для людей и домашних животных. Использование аденовирусного вектора сильно нацелено, поскольку оно индуцирует иммунитет при применении через слизистые оболочки.
Феномен иммунологической памяти широко используется в практике вакцинации людей для создания напряженного иммунитета и поддержания его длительное время на защитном уровне. Осуществляют это 2-3-кратными прививками при первичной вакцинации и периодическими повторными введениями вакцинного препарата - ревакцинациями .
Однако феномен иммунологической памяти имеет и отрицательные стороны . Например, повторная попытка трансплантировать уже однажды отторгнутую ткань вызывает быструю и бурную реакцию - криз отторжения.
В отличие от классических вакцин основной иммунный ответ не против вставленных генов, а против кодируемых ими белков. Этот процесс приводит к входу этих плазмид в клетки, смежные с местом инъекции. Иммунизация этим методом имеет некоторые необычные характеристики, например, реакция антитела протекает медленно, достигая пика только через 10 недель и, хотя и слабая, ответ очень длительный, и в экспериментах с морскими свинками этот ответ стал постоянным, Эта характеристика иммунизации в течение длительного периода времени является одним из основных преимуществ этого метода и вызывает большие надежды в научном и медицинском сообществе.
Иммунологическая толерантность -явление,противоположное иммунному ответу и иммунологической памяти.Проявляется она отсутствием специфического продуктивного иммунного ответа организма на антиген в связи с неспособностью его распознавания.
В отличие от иммуносупрессии иммунологическая толерантность предполагает изначальную ареактивность иммунокомпетентных клеток к определенному антигену.
Механизм действия этой вакцины очень мало известен. Что сделано до сих пор, так это сформулировать гипотезы о том, что происходит с помощью некоторых доказательств реакции организма. Это, как правило, вызывает аллергию - отсутствие косимуляторных сигналов - или неиммунный ответ - очень низкие уровни представления, которые мы видели, чего не происходит. Предлагаются две гипотезы, которые пытаются объяснить этот факт, но никто не смог утвердиться как истинный. Но эти клетки молчат и для начала процесса ответа потребуется стимул.
Признаки активации этих дендритных клеток плохо изучены. Другая проблема заключается в том, что дендритные клетки имеют ограниченный срок службы, что противоречит представлению и длительному иммунному ответу. Вторая гипотеза предполагает отложение антигенных комплексов и антител с низким сродством. При этом будет постоянное выделение нескольких антигенов, обеспечивающих длительный иммунный ответ.
Иммунологическую толерантность вызывают антигены, которые получили название толерогены. Ими могут быть практически все вещества, однако наибольшей толерогенностью обладают полисахариды.
Иммунологическая толерантность бывает врожденной и приобретенной. Примером врожденной толерантности является отсутствие реакции иммунной системы на свои собственные антигены. Приобретенную толерантность можно создать, вводя в
Несмотря на отсутствие знаний о механизме работы полинуклеотидной вакцины, существуют большие преимущества этого метода по сравнению с классическими вакцинами. Наиболее очевидным преимуществом является возможность манипулирования этими очень большими плазмидами. С помощью самых разных методов можно выбирать гены и модифицировать их. Другим преимуществом была бы высокая стабильность. Он также имеет большую характеристику отсутствия риска превращения в вирулентность. Его единственным недостатком является небольшая вероятность вставки этих генов в клеточный геном и вызывать онкогению.
организм вещества, подавляющие иммунитет (иммунодепрессанты), или же путем введения антигена в эмбриональном периоде или в первые дни после рождения индивидуума. Приобретенная толерантность может быть активной и пассивной. Активная толерантность создается путем введения в организм толерогена, который формирует специфическую толерантность. Пассивную толерантность можно вызвать веществами, тормозящими биосинтетическую или пролиферативную активность иммунокомпетентных клеток (антилимфоцитарная сыворотка, цитостатики и пр.).
В настоящее время в этой области проводится несколько исследований и разработок вакцин. Его исследование в основном нацелено на производство вакцин, вводимых перорально, для стимуляции иммунной системы, вызывающих гибель животного, и последующее изгнание нематоды из пищеварительного тракта . Это уменьшит или даже прекратит использование медицины против этих организмов.
На сегодняшний день он доступен только для ветеринарного использования. Другим большим преимуществом является то, что представление антигенов, продуцируемых для цитотоксических Т-лимфоцитов, вызывает клонирование экспрессии антиген-специфического, но оно способно распознавать гетерологичные линии, которые иммунизированы, тем самым защищая человека, иммунизированного против нескольких линий за один раз. Это не относится к антителам, которые являются «уникальными» для одной линии. Развитие этих новых вакцин на основе вирусов или рекомбинантных бактерий, пептидов и векторных плазмид обеспечивается недавними достижениями в области иммунологии, молекулярной биологии и биохимии пептидов.
Иммунологическая толерантность отличается специфичностью - она направлена к строго определенным антигенам. По степени распространенности различают поливалентную и расщепленную толерантность. Поливалентная толерантность возникает одновременно на все антигенные детерминанты, входящие в состав конкретного антигена. Для расщепленной, или моновалентной, толерантности характерна избирательная невосприимчивость каких-то отдельных антигенных детерминант.
Однако эти методы до сих пор не используются для массовой вакцинации, и большинство из них все еще проходят клинические испытания . Ни одна из этих различных вакцин, которые разрабатываются, уже не может быть полностью эффективной для профилактики инфекционных заболеваний или иммунотерапии против рака. Но преимущества и преимущества, которые они обещают, принесли большие надежды. Вирусные рекомбинантные вакцины, а также те, которые основаны на вакцинии или вирусе аденовируса, вызывают сильные иммунные ответы.
Вирус вакцины обладает тем преимуществом, что он довольно стабилен и иммуногенен при применении перорально, что делает его хорошим кандидатом для иммунизации диких животных. Рекомбинанты на основе дефектной репликации аденовируса более безопасны и также более эффективны по сравнению с рекомбинантами вирусной вакцины. Кроме того, они вызывают отличную иммунизацию при применении к слизистым оболочкам, что указывает на их использование в качестве вакцины против инфекционных агентов , поступающих в организм через дыхательные пути или половые пути.
Степень проявления иммунологической толерантности существенно зависит от ряда свойств макроорганизма и толерогена. Важное значение в индукции иммунологической толерантности имеют доза антигена и продолжительность его воздействия. Различают высокодозовую и низкодозовую толерантность. Высокодозовую толерантность вызывают введением больших количеств высококонцентрированного антигена. Низкодозовая толерантность, наоборот, вызывается очень малым количеством вы-сокогомогенного молекулярного антигена.
Пептиды все еще имеют ограниченные преимущества в профилактике инфекционных заболеваний, но они перспективны как вакцина в терапии рака. Пока безопасность и эффективность этих вакцин могут быть подтверждены, они могут принести иммунитет многочисленным патологическим агентам, тем самым улучшая стандарт и продолжительность жизни как людей, так и животных, жизненно важных для нашего выживания.
Является изучение ответов организма, которые обеспечивают иммунитет, то есть защиту от болезней. Хотя иммунная система очень сложна, некоторые компоненты иммунной системы легко обнаруживаются, например, антитела. Антигены - чужеродное вещество, которое индуцирует иммунный ответ, вызывая продуцирование антител и / или сенсибилизированных лимфоцитов, которые специфически реагируют с веществом; иммуноген.
Механизмы толерантности многообразны и до конца не расшифрованы.Известно,что ее основу составляют нормальныепроцессы регуляции иммунной системы. Выделяют три наиболее вероятные причины развития иммунологической толерантности:
Элиминация из организма антигенспецифических клонов лимфоцитов.
Блокада биологической активности им-мунокомпетентных клеток.
Антитело - белок сыворотки, который был индуцирован и специфически реагирует с посторонним веществом; иммуноглобулин. Эти антигены могут быть вирусами, клетками или молекулами белка. Иммунная система представляет собой сложную организацию биологически активных тканей, клеток, клеточных продуктов и медиаторов, и все они взаимодействуют для получения иммунного ответа. Иммунный ответ распознает и запоминает различные антигены. Специфический иммунитет характеризуется тремя свойствами.
Память специфики распознавания. Признание относится к способности иммунной системы распознавать различия в очень большом числе антигенов и различать их. Специфика относится к способности направлять ответ на определенный антиген. Память - это ссылка на способность иммунной системы запоминать антиген долго после первоначального контакта.
Быстрая нейтрализация антигена антителами.
Феномен иммунологической толерантности имеет большое практическое значение. Он используется для решения
многих важных проблем медицины, таких как пересадка органов и тканей, подавление аутоиммунных реакций , лечение аллергий и других патологических состояний, связанных с агрессивным поведением иммунной системы.
Основными тканями и органами иммунной системы являются. Они являются основными клетками, ответственными за иммунный ответ: Т-лимфоциты и В-лимфоциты. Периферические лимфоидные органы и ткани - лимфатические узлы , селезенка, кишечник-ассоциированной лимфоидной ткани , аппендикс, миндалины, пейеровы бляшки и лимфоидной ткани, связанные с бронхами.
Иммуноглобулины представляют собой белки, продуцируемые плазматическими клетками и секретируемые в организме в ответ на воздействие антигена. Это преобладающий иммуноглобулин в слезах, слюне, респираторных выделениях и желудочно-кишечном тракте . Обеспечивает защиту от организмов, которые вторгаются в эти районы.
№ 64 Классификация гиперчувствительности по Джейлу и Кумбсу.
Изучение молекулярных механизмов аллергии привело к созданию Джеллом и Кумбсом в 1968 г. новой классификации . В соответствии с ней различают четыре основных типа аллергии: анафилактический (I тип), цитотоксический (IIтип), иммунокомплексный (IIIтип) и опосредованный клетками (IV тип). Первые три типа относятся к ГНТ, четвертый - к ГЗТ. Ведущая роль в запуске ГНТ играют антитела (IgE, G и М), а ГЗТ - лимфоидно-макрофагальная реакция.
Иммунная система обладает двумя поистине удивительными свойствами : специфическим распознаванием и иммунной памятью. Под последней понимают способность развивать качественно и количественно более эффективный иммунный ответ при повторном контакте с тем же патогеном. Согласно этому различают первичный и вторичный иммунный ответ. Первичный иммунный ответ реализуется при первом контакте с незнакомым антигеном, а вторичный - при повторном. Вторичный иммунный ответ является более совершенным, так как осуществляется на качественно более высоком уровне из - за наличия преформированных иммунных факторов, отражающих генетическую адаптацию к патогену (уже имеются готовые гены специфических иммуноглобулинов и антиген - распознающих рецепторов Т - клеток) . Действительно, здоровые люди не болеют дважды многими инфекционными заболеваниями , так как при повторном заражении реализуется вторичный иммунный ответ, при котором отсутствует длительная воспалительная фаза, а в работу сразу же вступают иммунные факторы - специфические лимфоциты и антитела.
Вторичный иммунный ответ характеризуется следующими признаками:
1 . Более ранним развитием , иногда - даже молниеносным.
2 . Меньшей дозой антигена, необходимой для достижения оптимального иммунного ответа.
3 . Увеличением силы и продолжительности иммунного ответа за счёт более интенсивной продукции цитокинов (ТД 1 или че 2 профиля, в зависимости от природы патогена) .
4 . Усилением клеточных иммунных реакций за счёт более интенсивного образования специфических Т - хелперов 1 типа и цитотоксических Т - лимфоцитов.
5 . Усилением образования антител за счёт формирования большего количества Т - хелперов 2 типа и плазматических клеток.
6 . Повышением специфичности распознавания иммуногенных пептидов Т - лимфоцитами за счёт увеличения аффинности их антиген - специфических рецепторов.
7 . Повышением специфичности синтезируемых антител за счёт изначальной продукции IgG высокой аффинности / авидности.
Следует отметить, что невозможность формирования эффективной иммунной памяти является одним из характерных симптомов иммунодефицитных заболеваний человека. Так, у пациентов с гипоиммуноглобулинемией наблюдается феномен множественных эпизодов т.н. детских инфекций, так как после перенесённых инфекционных болезней не формируется защитный титр антител. Больные с дефектами клеточного иммунитета также не формируют иммунную память на Т - зависимые антигены, что проявляется отсутствием сероконверсии после инфекций и вакцинаций, однако общие концентрации иммуноглобулинов в их сыворотке крови могут быть нормальными.
Иммунологическая память - способность иммунной системы организма после первого взаимодействия с антигеном специфически отвечать на его повторное введение. Механизм, лежащий в основе иммунологической памяти, окончательно не установлен. Наряду со специфичностью, иммунологическая память -- важнейшее свойство иммунного ответа.
Позитивная иммунологическая память проявляется как ускоренный и усиленный специфический ответ на повторное введение антигена. При первичном гуморальном иммунном ответе после введения антигена проходит несколько дней (латентный период) до появления в крови антител. Затем наблюдается постепенное увеличение кол-ва антител до максимума с последующим снижением. При вторичном ответе на ту же дозу антигена латентный период сокращается, кривая увеличения антител становится круче и выше, а её снижение происходит медленнее. После стимуляции антигеном происходит пролиферация лимфоцитов (расширение клона), что приводит к образованию большого количества клеток исполнительного звена, а также других малых лимфоцитов, которые повторно входят в митотический цикл и служат для пополнения группы клеток, несущих соответствующий рецептор. Предполагается, что так как эти клетки результат вызванной антигеном пролиферации, то они способны к усиленному ответу при повторной встрече с антигеном (то есть, они действуют как клетки памяти). B семействе В-клеток эти клетки могут также подвергнуться переключению синтеза с IgM на IgG, что объясняет немедленное производство этими клетками IgG во время вторичного иммунного ответа.
Позитивная иммунологическая память к антигенным компонентам окружающей среды лежит в основе аллергических заболеваний, а к резус-антигену (возникает при резус-несовместимой беременности)-- в основе гемолитических болезни новорождённых.
Негативная иммунологическая память -- это естественная и приобретённая иммунологическая толерантность, проявляющаяся ослабленным ответом или его полным отсутствием как на первое, так и на повторное введение антигена. Нарушение негативной иммунологической памяти к собственным антигенам организма является патогенетическим механизмом некоторых аутоиммунных заболеваний.
Иммунологическая память представляет собой разновидность биологической памяти, принципиально отличающуюся от нейрологической (мозговой) памяти по способу её введения, уровню хранения и объёму информации. Иммунологическая память при ответе на разные антигены различна. Она может быть краткосрочной (дни, недели), долговременной (месяцы, годы) и пожизненной. Основные носители иммунологической памяти -- долгоживущие Т- и В-лимфоциты. Из других механизмов иммунологической памяти (кроме клеток памяти) определенное значение имеют иммунные комплексы, цитофильные антитела, а также блокирующие и антиидиотипичные антитела. Иммунологическая память можно перенести от иммунного донора неиммунному реципиенту, переливая живые лимфоциты или вводя лимфоцитарный экстракт, содержащий «фактор переноса» или иммунную РНК. Информационная ёмкость -- до 106--107 бит на организм. У позвоночных включается более 100 бит в сутки. В филогенезе иммунологическая память возникла одновременно с нейрологической памятью. Полной ёмкости иммунологическая память достигает у взрослых животных со зрелой иммунной системой (у новорождённых и старых особей она ослаблена).